

")
Publication History
Submitted: August 08, 2023
Accepted: August 18, 2023
Published: September 25, 2023
Identification
D-0087
Citation
Khaliqa Minhas, Muhammad Sufian, Lubna Rasheed & Umer Liaqat. Evaluation of Beauveria Bassiana and Metarhizium Anisopliae in integration with new chemistry insecticide against red cotton Bug (Dysdercus Koenigii) (Fabricius). Dinkum Journal of Natural & Scientific Innovations, 2(09):545-570.
Copyright
© 2023 DJNSI. All rights reserved
545-570
Evaluation of Beauveria Bassiana and Metarhizium Anisopliae in integration with new chemistry insecticide against red cotton Bug (Dysdercus Koenigii) (Fabricius)Original Article
Khaliqa Minhas ![]() 1*, Muhammad Sufian 2, Lubna Rasheed 3, Umer Liaqat 4
1*, Muhammad Sufian 2, Lubna Rasheed 3, Umer Liaqat 4
- Department of Zoology, Department of Zoology, University of Agriculture, Faisalabad, Pakistan; khaliqaminhas3@gmail.com
- Associate Professor; Department of Entomology, University of Agriculture, Faisalabad, Pakistan; muhammad.sufyan@uaf.edu.pk
- Department of Zoology, Department of Zoology, University of Agriculture, Faisalabad, Pakistan; lubnarasheed0@gmail.com
- Department of Entomology, University of Agriculture, Faisalabad, Pakistan; umeramer59@gmail.com
* Correspondence: khaliqaminhas3@gmail.com
Abstract: In the current study we used entomopathogenic fungus Beauveria bassiana and Metarhizium anisopliae in combination with the new chemical insecticide Movento and tested against red cotton bug, under lab conditions. The study conducted in the biological controlled lab under controlled conditions of temperature 25±2°C and 70±5% relative humidity using CRD layout in the Department of Entomology University of Agriculture Faisalabad. Different concentrations of fungi applied alone or in combination with recommended dose of Movento. Data was recorded on regular basis. The data obtained was subjected to ANOVA and treatment means with noticeable differences distinguished using LSD test p ≤ 0.05. The study will be useful for the sustainable management of red cotton bug in integration with other pest management strategies. The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (100%) was observed in case of integrated use of highest B. bassiana concentration. Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) The integrated applications of lower dose rate of. B. Bassiana and Movento caused higher nymphale mortality of D. koenigglei (90%) as compared to lower dose of M. anisopliae. The results of the experiment showed that maximum adult mortality of Dysdercus koeniggi (96.7%) was observed in case of integrated use of highest B. bassiana concentration. Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) The integrated applications of lower dose rate of. B. Bassiana and Movento caused higher adult mortality (83.3%) as compared to lower dose of M. anisopliae (Ma: 1x106conidia) (80%).ommended to be used as a teaching tool for faculty and students and a powerful power tool for local auto mechanic.
Keywords: beauveria bassiana, metarhizium anisopliae, insecticide, red cotton bug
- INTRODUCTION
Cotton industry and cotton derived products play a prominent role in the economic development of Pakistan and generate 63.9 percent of the overall agricultural exports [1]. It typically facilitates domestic cotton industry with raw material consisting of 1263 weaving factories, 503 textile mills, 2622 oil expelling machines and 8.1 million spindles. It also produces 3.5 to 3.6 million tons of cotton seeds, used for over 64% of the total domestic production of edible oil [2]. Cotton is primarily grown in the country’s provinces of Punjab and Sindh. The manufacturing method includes the fair usage of crop, nutrients, fertilizers and irrigation inputs. Cotton output has risen dramatically over the last decade but high yields have not yet been optimized. However, new processing technology and careful use of inputs at the accessible rates will improve cotton output. Various researches have been carried out to establish the factors responsible for improving productivity and thereby benefiting the farmers. They found that high input costs, untrained farmers, lack of market access, and financial capital shortages are responsible for reduction in per hectare yield and consequently reduced benefits for farmers. They estimated that the usage of inputs has a direct impact on the farmers’ productivity and benefit. He observed that the significant variables in cotton output are, irrigation, sowing cost, pesticide, crop, fertilizer, cultivation cost and labour. Plant safety and irrigation are the most significant variables that impact production costs [3]. Pakistan has two major crop seasons, namely the rabi and the kharif. Cotton, Maize, Rice, sugarcane, jowar and bajra are kharif crops while rabi crops are mustard, wheat, rapeseed, tobacco, barley and gram. Cotton is the largest non-food kharif crop and a largest foreign exchange income source. It constitutes 8.2 per cent of agricultural value added and around 2 percent of GDP [4]. Cotton also supplies feed, fiber, and edible oil. It supplies the raw materials for weaving factories, factories, and oil expellers [5]. Cotton is a major cash crop and support system of the cotton industry in Pakistan. During the “Kharif” or monsoon season from April to June, the cotton crop is planted on 15 per cent of arable land. Output is concentrated in two provinces with nearly 75 per cent of Punjab and nearly 25 per cent of Sindh region. Cotton is produced for the most part by small farmers who develop less than five hectares of land. Cotton expands to an approximate 1.6 million growers. The textile industry is Pakistan main manufacturing field, responsible for around 40 per cent of agricultural labour and hiring 10 million workers. The industry also produces 8% of Gross Domestic Product and more than 50% of export earnings, the highest of any other industry. The combined cotton and textile industries include 430 garment mills, 1050 ginneries, and 350 crushers and oil refiners [6]. It is very difficult for the farmers to deal with the noxious species – livestock pests (birds, snails, slugs, rats, nematodes, mites and insects), various plant diseases caused by (microbes, fungus, bacteria) and wild weeds, are collectively named pests. These pests are harmful for agricultural products cultivated for human consumption and utilization since the beginning of cultivation around 10,000 years ago. As with the abiotic causes of crop losses, especially the lack or abundance of water during the growing season, severe temperatures, high or low irradiance (factors that can only be managed within limited limits) and nutrient availability, biotic stressors have the capacity to significantly reduce crop output. This species may be managed by introducing physical (cultivation, mechanical weeding, etc.) and chemical steps (pesticides) to these species [7]. In Pakistan, the red cotton bug was considered as a minor pest but it has now turned to the major pest due to its cotton boll feeding habit with its sucking and piercing mouth parts. The major cause of cotton staining is believed to be the D. koenigii. In Pakistan during 2011 the cotton stainer played a vital role in destruction of cotton in the various cotton zones. From the last three years the population of the red cotton bug has been increased before 2011 the population size was small. The major attack is being done by red cotton bug in 2011 which resulted in the opened cotton bolls and stained lint [8]. Usually pests are controlled by using chemical pesticides but it has adverse effects on the environment, human health and non-targeted insects. Apart from these it causes resistance in the insect pest and the pesticide will no longer work against that pest [9-11]. Cotton stainer has been declared as most damaging cotton pests. Sprenkel, 2000 stated that not only the adults but nymphal stages also feed on the seed within the developing cotton bolls, resulting stained lint. So, this feeding habit of cotton stainer cause the reduction in size of cotton bolls, or the fruiting body which results in dropping of them on the ground [12]. In 2011 cotton stainer has caused serious losses in the major cotton growing areas of Pakistan. Badly opened bolls and stained lint was reported due to the severe attack of cotton stainer. However, little work is being done on this and a few research articles are being published on cotton stainer [13]. Traditionally pesticide have been used to control red cotton bug because is results in a quick knock down effect of pest as compared to other techniques. A variety of chemically derived insecticides are being used to control the insect pests but it has a variety of adverse effects on the environment and human health. So, in order to control these pests, we have to use other ecologically suitable and economically sound techniques [14]. Biological control can be used as an alternative strategy to insecticides which are harmful to the environment. The pathogenic fungi B. bassiana and M. anisopliae are diversely distributed and can be used to control the variety of insects and arachnids [15]. Fungal pathogenesis is a type of process in which the fungus enters in the insect host cuticle, damages it and cause infection which results in death of the insect pest [16]. The mechanism of pathogenesis of fungus includes the germination of spore, its penetration to the cuticle of insect pest and cause destruction of the insect biological systems which results in the mortality of the insect. The cuticle is the first line of defence, therefore direct attack of fungus on the cuticle is significant for virulence [17]. The insect pro-cuticle is primarily chitinous in nature with embedded proteins and its dispersion involves both enzymatic and mechanical components. The specificity of the fungal infection can be determined by the cuticular penetration and damage stage as many pathogens show their virulence when they are being injected into the host hemolymph. The fungal spore germinates with the formation of a thin germ tube which penetrates into the cuticle of insect via mechanical and enzymatic means [18]. The growing hyphae of fungus exerts mechanical pressure on the cuticle and releases organic acids to weaken the cuticular structure which results in the lipid breakdown of the cuticular membrane [19]. In M. anisopliae, the hypha formation, hydrophobins and the activation of destructive proteases is done by low nutrient level [20] where as in B. bassiana requires at least two hydrophobins (Hyd1 and Hyd2) which collectively cause cell surface hydrophobicity, adhesion and penetration to hydrophobic surfaces, and virulence. Keeping in view the importance of present study evaluation of M. anisopliae and B. bassiana in integration with the insecticide (movento) against the red cotton bug D. koenigii will be assessed in lab conditions.
- LITERATURE REVIEW
Roy and Pell [21] examined that natural enemies of pathogens and arthropods would contribute either as individual species or as complex species to the control of insect pest population. Natural enemies may interact either symbiotically or antagonistic. This paper reviewed the possible interactions between natural enemies of insects and fungi. In this article, it examines the possible relationship between insect and fungal natural enemies. In general, these studies show the positive nature of the relationships between natural enemies of arthropods and fungal pathogens in insect pest management. Omer et al. [22] explored the possibility of treating plants with natural elicitors for herbivore resistance. Applications of the natural plant inductor like Jasmonic acid (JA), were investigated in cotton plants for effects of induced resistance to cotton aphids Aphis gossypii, the western floral thrips Frankliniella occidentalis and spider mites Tetranychus urticae The preference for aphides and spider mites was reduced by more than 60 %, while for JA-induced leaves the preference was reduced by more than 90 % in thrips as compared to the controlled leaves. Aphid nymph’s survival was reduced by two-fold, and the percentage of the adult nymphs is eight-fold less than the controls for induced leaf. The reproduction and survival rate of adult aphids has decreased by 40 to 75% on induced leaf while the induction has no effect on thrips or spider mite’s survival. However, induction decreased thrip feeding by up to 80% and spider mites’ egg production by more than 75% and compared with controls. Inducing cotton seedlings decreased leaf area by approximately 28%. Oerke [23] investigated the productivity of crops cultivated for human consumption is at risk because of the occurrence of pests, pathogens, particularly weeds and animal pests. Crop losses incurred by these invasive species can be severe and crop conservation steps can be avoided or minimized by some precautionary measures. A summary of various crop losses and various pest control strategies established over the last century is presented in this study. In view of existing crop protection activities, estimates of possible and real losses for the period 2001-03 are provided regionally (19 regions) and overall, for cotton, wheat, rice, potatoes, soybean and maize. Between crops, the overall global possible losses in cotton production ranged from about 80% to over 50% wheat. The losses for soybean, wheat and cotton are expected to 26% to 29% and for maize, rice and potatoes 31%, 37%, and 40%. Weeds have caused the most possible loss (34%) in general, with less loss caused by animal pests and pathogens that is 18% and 16% respectively. In cash crops, the effectiveness of crop defense was higher than in food crops. The weed control system can be controlled manually or chemically, but the effectiveness of synthetic chemicals worldwide was substantially greater than that in the management of animal pests or diseases. Regional efficacy variations are identified. In the last 40 years, crop losses have not decreased substantially, despite a strong increase in use of pesticides. However, use of pesticides has allowed farmers to increase the crop production and to change their production system without retaining the increased loss due to an enhanced sensitivity to the harmful effects of pests. Uma Devi et al. [24] examined Beauveria bassiana, a fungus that has been used to biologically regulate many insect organisms, as a genetically distinct “population complex.” Being primarily asexual, mating tests in this complex species cannot be extended to delimit species. Genetic studies provide an effective way to classify isolated organisms. To this end, a molecular genetic analysis of a two samples of B. bassiana is performed. Among the Two subsample B. bassiana isolates, one of which represent a worldwide set and one of which belongs to a localized epizootic group. DNA markers were analyzed for different allelic variants of three genes (large and small subunits of rRNA and β-tubulin) through AFLPs (amplified polymorphisms in length of fragment) and SSCPs (single-strand conformation polymorphisms) and nucleotide series sequences. Within epizootic Community of B. bassiana, prevalence of recombination was evident from random combination of alleles in multi-locus tests and lack of phylogenetic concordance between 3 genealogies. The global study of B. bassiana is mainly cloned, with a divergence in species which leads to cryptic speciation, the recombination of isolates with a narrow ecological niche being common place. Brück et al. [25] researched that spirotetramat (Movento ® brand) tetramic acid derivative as excellence in laboratory / greenhouse studies as well as semi-field and field trials against sucking insect pests. The findings published show that the substance prevents the lipid biosynthesis and has more impacts on adult reproduction in the juvenile stages. No other insecticide is cross-resistant. Spirotetramat penetrates the leaf cuticle after its application in foliar transferring it through xylem and phloem in the form of spirotetramat-enol to rising shoots and roots. Spirotetramat has been developed in the Bayer Crop Science AG worldwide and has been introduced to various species, including white flies, mealy bugs, scales, psyllids, aphids and thrips in seeds, citrus trees, soya, pomace, wheat, grapes, puma and hops, bananas and stone fruits. The new mode of action makes Spirotetramat an outstanding revolving companion for established psyllide, white fly, aphid and treatment products that often are resistant to traditional insecticides. In comparison, the benefiting arthropods that make the substance ideal for current IPM systems have had only low adverse effects. These special properties help to preserve both the consistency and the quantity of the crop yield output. Compared to traditional insecticides and emerging technologies for IPM systems, modern chemistry is discussed in this report. Boldo et al. [26] examined the effect of chitinases in the remodeling of fungal cords and their role for nutrition and competition in exogenous Chitinase degradation. Because of the varied existence of these enzymes, unique roles are still assigned to every chitinase. Metarhizium anisopliae entomopathogenic fungus develops many chitinases, and here we determine if the pathogenicity of that fungus contains endochitinase CHI2. We developed strains that either exaggerate or ignore CHI2 chitinase. Southern, western and northern blot analyses and chitinase development confirmed these components. The Dysdercus peruvianus cotton stain bug was used as a host to control effects of CHI2 chitinase in virulence of the fungus. Higher efficiency in host killing shows that the development of this chitinase by a substitutionary supporter decreases the time required to destroy the bug. CHI2 overyex pressure buildings significantly, in contrast with humans, the knockout models displayed reduced virulence in insects. The absence of this single CHI2 chitinase reduced efficiency fungal infection but not any other detectable phenotype, indicating the M. anisopliae in insect infection, family 18, CHI2 Subgroup B plays a role. Biopesticides were tested with cotton leaves dipped into the respective treating solutions and air dried to remove excess water against Phenacoccus solenopsis and Paracocius marginatus adult nymphs and adults. Acephate recorded the highest mortality of 53.3 % at 48 hours following treatment among different therapies tested. The fish oil rosin soap, mealy quit and chlorpyrifos causes of 51.1 to 52.2 % mortality were equally poisonous. Further studies are underway to test the field results [27]. Agricultural pests, termites, and biological predators like mosquitoes and ticks have been widely used as chemical insecticides. The adverse effects on environmental and public health of synthetic chemical insecticides have led to comprehensive studies into alternatives. The Metarhizium fungal infection on terrestrial and aquatic larvae is illustrated in the review [28]. Beauveria bassiana as a biopesticide remedies were used against the Cimex lectularius bed bug. After one hour of treatment, the bugs are switched to unsprayed atmosphere and survival testing is carried out. In all bioassays immediate demise was found with bed bugs exposed to cotton knitted jersey die more rapidly [29].
- MATERIALS AND METHODS
The current experiment was carried out under the controlled conditions in Biological Control Lab of Entomology Department Faculty of Agriculture, University of Agriculture, and Faisalabad, Pakistan.
3.1 Insect material
The red cotton bug nymphs and adults were captured from different fields of Young Wala Fields, University of Agriculture, and Faisalabad as shown in Figure I. The insects we used in this assay were reared in the biological control lab of Department of Entomology in cages of size (30×30×30cm) and the environmental temperature maintained at 35°C while keeping 70-75% relative humidity.
3.2 Rearing of Red Cotton Bug
Preparing and administering an artificial diet on insects also poses several obstacles. Successive dietary proportion influences the insect biology and reproduction. In several biological studies, Dysdercus cingulatus [30], an economically significant insect pest infesting most of the Malvaceae species, has been used. Laboratory rearing on natural hosts is a time consuming, laborious and not economically viable one. An artificial diet has been established using its natural feed, the cotton seed, to resolve these constrain an artificial diet was established utilizing its natural feed, the cotton seed, to circumvent these constrains. D. cingulatus grew rapidly (17 days) with heavier female adults (148 mg/animals) and optimum fecundity (57 eggs/person) while reared with artificial diet. Artificial food, however, decreases the risk of nymphal survival (10 %), adult lifespan (3 and 2 days for male and female respectively) and gender-biased sex. Additionally, chemical food significantly improves hatchability. The pest’s oviposition index and relative growth rate may be attributed to the higher starch, protein and lipid content found in the artificial diet. Therefore, it is indicated that the current experimental diet could be used for experimental pest rearing.

Figure 1: Rearing of Red Cotton Bug
3.3 Nymphs:
Five nymphal stages or instars are available. Typically, the first one is spent underground. Typically, the length of any of the first four phases during the midsummer ranges four to five days, although the fifth stage normally takes almost twice as long. All five phases include a full production cycle from 21 to 35 days (Sprenkel, 2000). Overall, the nymphs are red. The fourth and fifth in stars have dark wing surfaces, and when maturity progresses, the distinguishing lines between abdominal segments become very distinct.

Figure 2: Red cotton bug hatching

Figure 3: Red cotton bug on cotton plant

Figure 4: Red cotton bug destroying cotton boll

Figure 5: Eggs of Red cotton bug eggs
3.4 Experimental Layout
In total 10 specimens per replication were immersed in various concentrations of entomopathogenic fungi and new chemistry pesticide movento was used. Three treatments including control and each treatment were replicated three times. Adults of red cotton bug were individually sprayed with the fungus solution and pesticide for approximately 8-10 seconds. The immersed insects were dried out by placing them on the tissue paper so that they soak the excess moisture, shifted into the plastic boxes with the cotton bolls as feed. Mortality data was recorded on regular basis and cadavers were placed in disinfected petri plates with wet cotton pads to for the sporulation of fungus.

Figure 6: Cages in which the experiment was conducted

Figure 7: Red cotton bugs present in cages

Figure 8: Different concentration of fungus used in the experiment
3.5 Entomopathogenic fungi
B. bassiana (Racer) and M. anisopliae (pacer) product was obtained from Agri. Life Hyderabad India and used against red cotton bug. For the management of red cotton bug different concentration of the fungus were applied. High humidity was required for the optimal activity of fungus. Application of entomopathogenic fungi was efficient for the managing of pest population during their initial stages of growth. Mortality of insect depends on viability and number of spores contacting the pest.
3.6 Mycosis and Sporulation
For sporulation and mycosis, the sterile petri plates were used to place the cadavers of fungal treated C. patellus after that they are transferred to the plastic vials and refrigerated at 4°C. Sodium hypochlorite solution (0.05%) was used to sterilize the collected cadavers followed by the distilled water wash. For the external fungal growth, the cadavers were cultivated on PDA plates and incubated at 75±5% R. H and 25± 1 °C temperature for 1 week. The cadaver then placed under the microscope for sporulation determination. The cadavers duplication was combined with a drop of Tween-80 in 20ml distilled water and stirred for 10 min to assess the sporulation. A hemocytometer under a microscope was used to measure the total number of conidia /ml [27].
3.7 Statistical Analysis
The data gathered was subjected to analysis of variance (ANOVA) and treatment means with noticeable differences separated using LSD test p ≤ 0.05. All statistical analysis was carried out using Statistics 8.1 pakages.
- ANALYSIS AND RESULTS
4.1. Nymphal mortality after day 1
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the nymphal mortality of Dysdercus koeniggi after one day of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 1). Overall, the results showed that integrated applications of tested treatments showed more pathogenicity towards the nymphal instar of D. koeniggi nymphs as compared to their individual applications.
Table 1: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after one day of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 6936.67 | 770.74 | 11.01 | < 0.001 |
| Error | 20 | 1400 | 70 | ||
| Total | 29 | 8336.67 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
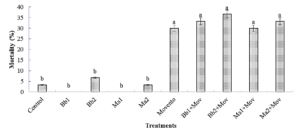
Figure 9: Effect of different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) on nymphal mortality of Dysdercus koeniggi after day 1 of treatments application. Bars with different lower-case letters are significantly different (LSD test at p ≤ 0.05).
Where; Bb1: 1×106 conidia/ml; Bb2: 1×108 conidia/ml; Ma1: 1×106 conidia/ml; MA2: 1×108 conidia/ml; Mov: Movento: (1%) (240 SC).
4.2. Nymphal mortality after day 2
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the nymphal mortality of Dysdercus koeniggi after two days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 10). Overall, the results showed that integrated applications of tested treatments showed more pathogenicity towards the nymphal instar of D. koeniggi nymphs as compared to their individual applications. Moreover, a significantly higher mortality was reported in case of higher fungal concentrations as compared to their lower doses. The results further revealed that B. bassiana showed more pathogenicity towards nymphal instar of D. koeniggi as compared to M. anisopliae (Figure 10).
Table 2: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after two days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 16520 | 1835.6 | 17.76 | < 0.001 |
| Error | 20 | 2066.7 | 103.3 | ||
| Total | 29 | 18586.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.3. Nymphal mortality after day 3
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the nymphal mortality of Dysdercus koeniggi after three days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 3). The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (80%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) followed by the integration of highest concentration of M. anisopliae (Ma2: 1×108 conidia/ml) and Movento (1%). Similarly, the integrated applications of lower dose rate of B. bassiana (Bb1: 1×106 conidia/ml) and Movento (1%) also caused higher nymphal mortality of D. koeniggi (73.3%) as compared to combined treatment of lower dose of M. anisopliae (Ma1: 1×106 conidia/ml) and Movento (1%) (70%).
Table 3: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after three days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 27030 | 3003.3 | 30.03 | < 0.001 |
| Error | 20 | 2000 | 100 | ||
| Total | 29 | 29030 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.4. Nymphal mortality after day 4
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the nymphal mortality of Dysdercus koeniggi after four days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 4).
Table 4: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after four days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 18136.7 | 2015.2 | 16.79 | < 0.001 |
| Error | 20 | 2400 | 120 | ||
| Total | 29 | 20536.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.5. Nymphal mortality after day 5
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the nymphal mortality of Dysdercus koeniggi after five days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 5).
Table 5: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after five days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 16230 | 1803.33 | 25.76 | < 0.001 |
| Error | 20 | 1400 | 70 | ||
| Total | 29 | 17630 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.6. Nymphal mortality after day 6
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the nymphal mortality of Dysdercus koeniggi after six days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 4.6).
Table 6: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after six days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 17336.7 | 1926.3 | 13.13 | < 0.001 |
| Error | 20 | 2933.3 | 146.7 | ||
| Total | 29 | 20270 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.7. Nymphal mortality after day 7
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the nymphal mortality of Dysdercus koeniggi after seven days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi (Table 7).
Table 7: Analysis of variance table for nymphal mortality of Dysdercus koeniggi after seven days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 17533.3 | 1948.1 | 18.26 | < 0.001 |
| Error | 20 | 2133.3 | 106.7 | ||
| Total | 29 | 19666.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.8. Mycosis in nymphs
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the mycosis of nymphal cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the mycosis of nymphal cadavers of Dysdercus koeniggi (Table 8).
Table 8: Analysis of variance table for mycosis in nymphal cadavers of Dysdercus koeniggi subjected to different treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 30313.14 | 3368.13 | 111.26 | < 0.001 |
| Error | 20 | 605.47 | 30.27 | ||
| Total | 29 | 30918.6 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.9. Sporulation in nymphs
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the sporulation of nymphal cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the sporulation of nymphal cadavers of Dysdercus koeniggi (Table 9).
Table 9: Analysis of variance table for sporulation in nymphal cadavers of Dysdercus koeniggi subjected to different treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 107648.3 | 11960.9 | 79.19 | < 0.001 |
| Error | 20 | 3020.7 | 151 | ||
| Total | 29 | 110669 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.10. Adult mortality after day 1
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the adult mortality of Dysdercus koeniggi after one day of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 10).
Table 10: Analysis of variance table for adult mortality of Dysdercus koeniggi after one day of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 9146.7 | 1016.3 | 6.93 | < 0.001 |
| Error | 20 | 2933.3 | 146.7 | ||
| Total | 29 | 12080 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.11. Adult mortality after day 2
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the adult mortality of Dysdercus koeniggi after two days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 11).
Table 11: Analysis of variance table for adult mortality of Dysdercus koeniggi after two days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 20653.3 | 2294.8 | 15.65 | < 0.001 |
| Error | 20 | 2933.3 | 146.7 | ||
| Total | 29 | 23586.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.12. Adult mortality after day 3
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the adult mortality of Dysdercus koeniggi after three days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 12).
Table 12: Analysis of variance table for adult mortality of Dysdercus koeniggi after three days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 25896.7 | 2877.4 | 23.98 | < 0.001 |
| Error | 20 | 2400 | 120 | ||
| Total | 29 | 28296.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.13. Adult mortality after day 4
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the adult mortality of Dysdercus koeniggi after four days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 13).
Table 13: Analysis of variance table for adult mortality of Dysdercus koeniggi after four days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 19880 | 2208.9 | 11.04 | < 0.001 |
| Error | 20 | 4000 | 200 | ||
| Total | 29 | 23880 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.14. Adult mortality after day 5
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the adult mortality of Dysdercus koeniggi after five days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 14).
Table 14: Analysis of variance table for adult mortality of Dysdercus koeniggi after five days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 15030 | 1670 | 12.53 | < 0.001 |
| Error | 20 | 2666.7 | 133.3 | ||
| Total | 29 | 17696.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.15. Adult mortality after day 6
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the adult mortality of Dysdercus koeniggi after six days when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 15).
Table 15: Analysis of variance table for adult mortality of Dysdercus koeniggi after six days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 14070 | 1563.3 | 12.68 | < 0.001 |
| Error | 20 | 2466.7 | 123.3 | ||
| Total | 29 | 16536.7 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.16. Adult mortality after day 7
Analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the adult mortality of Dysdercus koeniggi after seven days of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi (Table 16). The results of the experiment showed that maximum adult mortality of Dysdercus koeniggi (96.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) followed by the integration of highest concentration of M. anisopliae (Ma2: 1×108 conidia/ml) and Movento (1%) (86.7%). Similarly, the integrated applications of lower dose rate of B. bassiana (Bb1: 1×106 conidia/ml) and Movento (1%) also caused higher adult mortality of D. koeniggi (83.3%) as compared to combined treatment of lower dose of M. anisopliae (Ma1: 1×106 conidia/ml) and Movento (1%).
Table 16: Analysis of variance table for adult mortality of Dysdercus koeniggi after seven days of treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 14296.67 | 1588.52 | 17.65 | < 0.001 |
| Error | 20 | 1800 | 90 | ||
| Total | 29 | 16096.67 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.17. Mycosis in adults
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the mycosis of adult cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the mycosis of adult cadavers of Dysdercus koeniggi (Table 17).
Table 17: Analysis of variance table for mycosis in adult cadavers of Dysdercus koeniggi subjected to different treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 22204.34 | 2467.15 | 47.78 | < 0.001 |
| Error | 20 | 1032.73 | 51.64 | ||
| Total | 29 | 23237.07 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
4.18. Sporulation in adults
Analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the sporulation of adult cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the sporulation of adult cadavers of Dysdercus koeniggi (Table 18).
Table 18: Analysis of variance table for sporulation in adult cadavers of Dysdercus koeniggi subjected to different treatments application
| SOV | DF | SS | MS | VR | P-Value |
| Treatment | 9 | 106898.7 | 11877.6 | 63.44 | < 0.001 |
| Error | 20 | 3744.4 | 187.2 | ||
| Total | 29 | 110643.1 |
SOV = Source of variation; DF = Degree of freedom; SS = Sum of Squares; MS = Mean Squares; VR = Variance Ratio; p ≤ 0.001 = Highly-significant
- DISCUSSION
Chemical Insecticides were found better against nymphs and adults in comparison with the biopesticides, nymphs were more vulnerable than adults. After 48h of treatment application acephate reported maximum mortality of 57.78 % adult followed by the Chlorpyriphos which showed a death rate of 55.56% of nymphs. At 48 hours after surgery, Verticillium lecanii reported more than 50% nymph death. Acephate showed maximum mortality with 55.56% when it was assessed on adults. All biologically regulated agents demonstrated hyphal penetration in the body of both the nymph and the adults. The combination of biopesticides, viz, B. bassiana, M. anisopliae V. lecanii, and with acephate has demonstrated significant nymph and adult deaths. In the current study on mealy bug showed the development of fungi on the insect body, cuticular degradation was followed. The secretion of hydrolytic enzymes can result in the cuticular breakdown [29-35]. The residual spray of B. bassiana showed rapid mortality in bed bugs that is within 3-5 days. The level of infection generally showed a 100% susceptibility under such exposures to fungus. The few individuals did not die on the first exposure but the second exposure to the fungal spray resulted in mortality of these few survivors. which indicates that spores have been sub optimally picked up rather than some physiological resistance (especially from paper substrate). Effects from six different experiments were stable [36]. All the chemically synthetic insecticides when applied on plants moved in an acropetal fashion that is in a one-way direction towards the shoot. But with the newly synthesized chemical insecticide that is movento the case is entirely different [37-56]. When spirotetramat penetrated to the plant phloem it is transformed into enol which travels both ways that is roots as well as shoots. It can control the broad spectrum of sucking insect pests some of the organisms can only be handled in field condition and difficult to treat them in laboratory conditions. Spirotetramat demonstrated outstanding effectiveness in a leaf-dip bioassay against 3-4-day cotton aphid nymphs [38]. In the present study (ANOVA) revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the nymphal mortality of Dysdercus koeniggi after 1st to 7th day of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the nymphal mortality of Dysdercus koeniggi. In the present study analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the mycosis of nymphal cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the mycosis of nymphal cadavers of Dysdercus koeniggi. In the experiment conducted analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the sporulation of nymphal cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the sporulation of nymphal cadavers of Dysdercus koeniggi. In the experiment conducted analysis of variance table revealed that the individual and integrated applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov) had a significant effect (p ≤ 0.05) on the adult mortality of Dysdercus koeniggi after 1st to 7th day of treatment application. The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the adult mortality of Dysdercus koeniggi.In the experiment conducted analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the mycosis of adult cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the mycosis of adult cadavers of Dysdercus koeniggi. In the experiment conducted analysis of variance table revealed that significant differences (p ≤ 0.05) were detected in the sporulation of adult cadavers of Dysdercus koeniggi when tested against different individual and combined applications of Beauveria bassiana (Bb), Metarhizium anisopliae (Ma) and Movento (Mov). The tested treatments differed significantly (p ≤ 0.05) from untreated control regarding the sporulation of adult cadavers of Dysdercus koeniggi. Overall, the results showed that integrated applications of tested treatments showed more pathogenicity towards the nymphal instar of D. koeniggi nymphs as compared to individual applications. The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (36.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC). The results of the experiment showed that maximum adult mortality of Dysdercus koeniggi (96.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) The integrated applications of lower dose rate of. B. Bassiana and Movento caused higher adult mortality (83.3%) as compared to lower dose of M. anisopliae (Ma: 1×106 conidia) (80%). The results of the experiment showed that maximum mycosis (91.2%) was observed in case of individual application of lowest concentration of B. bassiana treatments as compared to M. anisopliae treatments. A significantly higher Mycosis was reported in case. of B. bassiana treatments compared to M. anisopliae. Overall, the results showed that mycosis was more profound in. case of lower individual concentrations of fungal treatments as vs. their integrated applications. The results of the experiment showed that maximum sporulation (170.4 conidia/ml) was observed in case of individual application of lowest concentration of B. bassiana treatments as compared to M. anisopliae treatments. Results showed that mycosis was more profound in. case of lower individual concentrations of fungal treatments as compared to their integrated applications.
- CONCLUSION
The red cotton bug (Dysdercus koenigii F.) is considered a serious cotton pest particularly after boll opening. Adults and nymphs decrease plant productivity and crop yield by sucking cell sap from seeds. Traditionally, it has been controlled by synthetic insecticides that resulted in high residual effects and natural enemies’ disturbance. As an alternate the use of biocontrol agents that is entomopathogenic fungus can be a potential management technique. The study will be conducted in the biological controlled lab under controlled conditions of temperature 25±2°C and 70±5% relative humidity. Overall, the results of the study showed that integrated applications of tested treatments showed more pathogenicity towards the nymphal instar of D. koeniggi nymphs as compared to individual applications. The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (36.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC). The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (56.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) This compared to combined treatment of lower dose of M. anisopliae (Ma1: 1×106 conidia) (50%) and Moventos (1×106) ( 50%) Overall, the results showed that integrated applications of tested treatments showed more pathogenicity towards the nymphs as compared to their individual applications. The results of the experiment showed that maximum nymphal mortality of Dysdercus koeniggi (100%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) The integrated applications of lower dose rate of. B. Bassiana and Movento caused higher nymphal mortality of D. koenigglei (90%) as compared to lower dose of M. anisopliae. The results of the experiment showed that maximum adult mortality of Dysdercus koeniggi (96.7%) was observed in case of integrated use of highest B. bassiana concentration. (Bb2: 1×108 conidia/ml) and Movento (1%) (Mov: 240 SC) The integrated applications of lower dose rate of. B. Bassiana and Movento caused higher adult mortality (83.3%) as compared to lower dose of M. anisopliae (Ma: 1×106 conidia) (80%). The results of the experiment showed that maximum mycosis (91.2%) was observed in case of individual application of lowest concentration of B. bassiana treatments as compared to M. anisopliae treatments. A significantly higher Mycosis was reported in case. of B. bassiana treatments compared to M. anisopliae. Overall, the results showed that mycosis was more profound in. case of lower individual concentrations of fungal treatments as vs. their integrated applications. The results of the experiment showed that maximum sporulation (170.4 conidia/ml) was observed in case of individual application of lowest concentration of B. bassiana treatments as compared to M. anisopliae treatments. Results showed that mycosis was more profound in. case of lower individual concentrations of fungal treatments as. compared to their integrated applications.
REFERENCES
- Ahmed, K., S. Freed, R.F. Shoukat and K.W. Ahmad. 2020. Efficacy of Entomopathogenic Fungi with Insecticides Mixtures against Oxycarenus hyalinipennis (Costa) (Lygaeidae: Hemiptera). Pak. J. Zool., 52: 573-583.
- Ahmed, R., I. Nadeem, M.J. Yousaf, T. Niaz, A. Ali and Z. Ullah. 2015. Impact of dusky cotton bug (Oxycarenus laetus Kirby) on seed germination, lint color and seed weight in cotton crop. J. Entomol. Zool. Stud., 3: 335-338.
- Aw, K.M.S., S.M. Hue, M. Rafiq, et al. 2014. Evaluation of Bio-Pesticides Against Red Cotton Bug and Fruit Borer of N Save Nature to Survive, 5: 601-604.
- Aw, K.M.S. and S.M. Hue. 2017. Mode of infection of Metarhizium spp. fungus and their potential as biological control agents. J. Fungi., 3: 30-35.
- Banu, J. G., T. Surulivelu, M. Amutha and N. Gopalakrishnan. 2010. Laboratory evaluation of insecticides and biopesticides against Phenococcus solenopsis and Paracoccus marginatus infesting cotton. J. Biopestic., 3: 343-346.
- Barbarin, A.M., N.E. Jenkins, E.G. Rajotte and M.B. Thomas. 2012. A preliminary evaluation of the potential of Beauveria bassiana for bed bug control. J. Invertebr. , 111: 82–85.
- Boldo, J.T., A. Junges, K.B. Amaral, C.C. Staats, M.H. Vainstein and Schrank. 2009. Endochitinase CHI2 of the biocontrol fungus Metarhizium anisopliae affects its virulence toward the cotton stainer bug Dysdercus peruvianus. Curr. Genet., 55: 551- 560.
- , K., I. Hassan and A. Maqbool. 2005. Factors Affecting Cotton Yield: A Case Study of Sargodha (Pakistan). J. Agric. Soc. Sci., 1: 332-334.
- Beys-Da-Silva, W.O., L. Santi, M. Berger, D. Calzolari, D.O. Passos, J.A. Guimarães, J.J. Moresco and J.R.Yates. 2014. Secretome of the biocontrol agent Metarhizium anisopliae induced by the cuticle of the cotton pest Dysdercus peruvianus reveals new insights into infection. J. Proteome Res., 13: 2282-2296.
- Brück, E., A. Elbert, R. Fischer, S. Krueger, J. Kühnhold, A.M. Klueken, R. Nauen, J.F. Niebes, U. Reckmann, H.J. Schnorbach, R. Steffens and X.V. Waetermeulen. Movento®, an innovative ambimobile insecticide for sucking insect pest control in agriculture: Biological profile and field performance. J. Crop Prot., 28 :838- 844.
- Dar, S.A., B.A. Rather and A.A. Kandoo. 2017. Insect pest management by entomopathogenic J. Entomol. Zool. Stud., 5: 1185-1190.
- Egbuta, M.A., S.D.L.E. McIntosh, T.V. Waters, T. Vancov and L. Liu. 2017. Biological importance of cotton by-products relative to chemical constituents of the cotton plant. Molecules, 22: 12-14.
- El-Sharabasy, H.M. 2015. Laboratory evaluation of the effect of the entomopathogenic fungi, Thompsonii and Paecilomyces fumosoroseus, against the citrus brown mite, Eutetranychus orientalis (Acari: Tetranychidae). Plant Prot. Sci., 51: 39-40.
- González-Mas, N., M. Cuenca-Medina, F. Gutiérrez-Sánchez and E. Quesada-Moraga. 2019. Bottom-up effects of endophytic Beauveria bassiana on multitrophic interactions between the cotton aphid, Aphis gossypii, and its natural enemies in melon. J. Pest Sci., 92: 1271-1281.
- Hegedus, D.D. and G.G. Khachatourians. 1995. The impact of biotechnology on hyphomycetous fungal insect biocontrol agents. Biotechnol. Adv., 3: 455- 490.
- Howard, A.F.V., C.J.M. Koenraadt, M. Farenhorst, B.G.J. Knols and W. Takken. 2010. Pyrethroid resistance in Anopheles gambiae leads to increased susceptibility to the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana. Malar. J., 9: 1-9.
- Huang, Z., S. Ali, S. Ren, J. Wu and Y.Zhang .2012 . Influence of the entomopathogenic fungus Beauveria bassiana on Prynocaria congener (Billberg) (Coleoptera: Coccinellidae) under laboratory conditions. Pak. J. Zool., 44: 209-216.
- Husnain, S. 2014. Entomopathogenic fungi as potent agents of biological control. Int. J. Res. Technol., 2: 234-237.
- Heppner, J.B, J.L. Capinera, J. Ellis, A.N. Alekseev, P.G. Weintraub, J.L. Capinera, G. Manley, J.O. Schmidt, D.B. Rivers, D. Rivers, J.A. Hogsette, K.W. Stewart, P.A. Weston, R. Malakar-Kuenen, K. .M Daane, R.J. Prokopy, J . Sivinski, S.R. Wing and P.D. Shirk . 2008 . Verticillium lecanii. Ency. Entomol., 1:4095-4095.
- Irshad, M., M.M. Saleem and A. Hanif. 2019. Comparative efficacy of different insecticides against dusky cotton bug (Oxycarenus spp.) under field conditions. J. Entomol. Zool. , 7: 125-128.
- Jacobsen, S.K., I. Klingen, J. Eilenberg, B. Markussen and L. Sigsgaard. 2019. Entomopathogenic fungal conidia marginally affect the behavior of the predators Orius majusculus (Hemiptera: Anthocoridae) and Phytoseiulus persimilis (Acari: Phytoseiidae) foraging for healthy Tetranychus urticae (Acari: Tetranychidae). Appl. Acarol., 79 :299-307.
- Jaleel, W., S. Saeed and M.N. Naqqash. 2013. Biology and bionomics of Dysdercus koenigii (Hemiptera: Pyrrhocoridae) under laboratory conditions. Pak. J. Agric. Sci., 50: 373- 378.
- Khan, B.A., S. Freed, J. Zafar and M. Farooq. 2014. Evaluation of three different insect pathogenic fungi for the control of Dysdercus koenigii and Oxycarenus Pak. J. Zool., 46: 1759-1766.
- Kirkland, B.H., A. Eisa and N.O. Keyhani. 2005. Oxalic acid as a fungal acaracidal virulence J. Med. Entomol., 42: 346-351.
- Kodandaram, M.H., N.S.A. Thakur and A.N. Shy Lesha. 2008. Toxicity and morphogenetic effects of different botanicals on red cotton bug Dysdercus koenigii Fab. (Hemiptera: Pyrrhocoridae) in North Eastern Hill (NEH) region of India. Biopestic., 1:187-189.
- Leger, R.S., R.M. Cooper and A.K. Charnley. 1991. Characterization of chitinase and chitobiase produced by the entomopathogenic fungus Metarhizium anisopliae. J. Pathol., 58: 415-426.
- Meyling, N.V. and J. Eilenberg. 2007. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol Control., 43 :145-155.
- Mohyuddin, A.I., G. Jilani, A.G. Khan, A. Hamza, I. Ahmed and Z. Mahmood. 1997. integrated pest management of major cotton pests by conservation, redistribution and augmentation of natural enemies. Pak. J. Zool., 29: 293-298.
- Moorthi, P.V., C. Balasubramanian, P.B. Avery, A.N. Banu and T. Kubendran. 2012. Efficacy of fungal entomopathogens against red cotton stainer, Dysdercus cingulatus Fabricius (Hemiptera: Pyrrhocoridae). J. Biopest., 5: 140-145.
- Nasir, M., M.U. Asif and R.M. Shamraiz. 2019. Comparative efficacy of different insecticides against dusky cotton bug (Oxycarenus spp.) under field conditions. J. Entomol. Stud., 7: 125-128.
- Nauen, R., U. Reckmann, J. Thomzik and W. Thielert. 2007. Biological profile of spirotetramat (Movento)-a new two-way systemic (amimobile) insecticide against sucking pests Studies on the susceptibility of Tuta absoluta to novel and diamide insecticides View project Biological profile of spirotetramat Movento ® Bayer. Crop Sci., 61: 245-278.
- Oerke, E. C. 2006. Crop losses to pests. J. Agric. Sci., 144: 31-43.
- Omer, A.D., J. Granett, R. Karban and E.M. Villa. 2001. Chemically-induced resistance against multiple pests in cotton. Int. J. Pest Manage., 47: 49-54.
- Paula, A.R., A.T Carolino, C.O. Paula and R.I. Samuels. 2011. The combination of the entomopathogenic fungus Metarhizium anisopliae with the insecticide Imidacloprid increases virulence against the dengue vector Aedes aegypti (Diptera: Culicidae). Parasites Vectors, 4: 1-8.
- Riasat, T., W. Wakil, M. Ashfaq and S.T. Sahi. 2011. Effect of Beauveria bassiana mixed with diatomaceous earth on mortality, mycosis and sporulation of Rhyzopertha dominica on stored wheat. Phytoparasitica, 39: 325-331.
- Rafiq, M., S.I.A. Shah, M.T. Jan, I.R. Khan, S.A.S. Shah and Z. Hussain. 2014. Efficacy of Different Groups of Insecticides Against Cotton Stainer (Dysdercus Koenigii) in Field Pak. Entomol., 36: 105-110.
- Roberts, D.W. and R.J. St Leger. 2004. Metarhizium spp., cosmopolitan insect-pathogenic fungi: mycological aspects. Adv. Appl. Microbiol., 54: 1-70.
- Roy, H. E. and J. K. Pell. 2000. Interactions between entomopathogenic fungi and other natural enemies: Implications for biological control. Biocontrol Sci. Techn., 10: 737-
- Sahayaraj, K., M. Tomson and S. Kalidas. 2012. Artificial rearing of the red cotton bug, Dysdercus cingulatus using cotton seed-based artificial diet (Hemiptera: Pyrrhocoridae). Entomol. Gen., 33:283-288.
- Santi, L., L.A.D. e Silva, W.O.B. da Silva, A.P.F. Corrêa, D.E.N. Rangel, C.R. Carlini, A. Schrank and M. H. Vainstein. 2011. Virulence of the entomopathogenic fungus Metarhizium anisopliae using soybean oil formulation for control of the cotton stainer bug, Dysdercus peruvianus. World J. Microb. Biot., 27: 2297-2303.
- Sarwar, Z.M., M. Ijaz, M.A. Sabri, H. Yousaf. and M. Mohsan. 2018. Effects of selected synthetic insecticides on the total and differential populations of circulating haemocytes in adults of the red cotton stainer bug Dysdercus koenigii (Fabricius) (Hemiptera: Pyrrhocoridae). Environ. Sci. Pollut. Res. Int., 25:17033-17037.
- Schrank, A., and M.H. Vainstein .2010. Metarhizium anisopliae enzymes and toxins. Toxicon, 56 :1267-1274.
- Shah, S.I.A., M. Rafiq, T.H. Malik, I.R. Khan, S.A.S. Shah and Z. Hussain .2016. Comparison of the newly introduced rearing methods of cotton stainer, Dysdercus koenigii (hemiptera: Pyrrhocoridae) with classical methods. Pak. J. Zool., 48: 781-
- Shah, S.I.A. 2014. The cotton stainer (Dysdercus koenigii): An emerging serious threat for cotton crop in Pakistan. Pak. J. Zool., 46: 329-335.
- Sharif, I. and M. Bashir .2010. Munich Personal RePEc Archive Factors Affecting Cotton Production in Pakistan: Empirical Evidence from Multan District. 22829.
- Sprenkel, R.K. 2000. Cotton plant and pest monitoring manual for Florida.
- Sufyan, M., A. Abbasi, W. Wakil, M. D. Gogi, M. Arshad, A. Nawaz and Z. Shabbir. 2019. Efficacy of Beauveria Bassiana and Bacillus Thuringiensis Against Maize Stem Borer Chilo Partellus (Swinhoe) (Lepidoptera: Pyralidae). Gesunde Pflanz., 71: 197-204.
- Thomas, M.B. and A.F. Read. Can fungal biopesticides control malaria? Nat. Rev. Microbiol., 5: 377-383.
- Tiago, P. V., N.T. Oliveira and E.Á. de L.A. Lima. 2014. Biological insect control using Metarhizium anisopliae: morphological, molecular, and ecological aspects. Cienc. Rural, 44: 645–651.
- Uma Devi, K., A. Reineke, N. Nageswara, C. Maheswara, J. 2006. Genetic diversity, reproductive biology, and speciation in the entomopathogenic fungus Beauveria bassiana (Balsamo) Vuillemin. Genome, 49: 495-504.
- Vu, V.H., S. Hong and K. Kim. 2007. Selection of Entomopathogenic Fungi for Aphid J. Biosci. Bioeng., 104: 498-505.
- Williams, D. (2015). Pakistan Cotton and Products Annual.
- Xiao, G., S.H. Ying, P. Zheng, Z.L. Wang, S. Zhang, X.Q. Xie, Y. Shang, R.J.S. Leger, G.P. Zhao, C. Wang and M.G. Feng. 2012. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Scientific Reports, 2: 483-489.
- Zacharuk, R.Y. 1971. Fine structure of the fungus Metarrhizium anisopliae infecting three species of larval Elateridae (Coleoptera). IV. Development within the host. Can. J. , 17: 25-529.
- Zhang, S., E. Widemann, G. Bernard, A. Lesot, F. Pinot, N. Pedrini and N.O. Keyhani. 2012. CYP52X1, representing new cytochrome P450 subfamily, displays fatty acid hydroxylase activity and contributes to virulence and growth on insect cuticular substrates in entomopathogenic fungus Beauveria bassiana. J. Biol. Chem., 287: 13477-
- Zheng, P., Y. Xia, G. Xiao, C. Xiong, X. Hu, S. Zhang, H. Zheng, Y. Huang, Y. Zhou, S. Wang and G.P. Zhao. 2012. Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biol., 12: R116.
Publication History
Submitted: August 08, 2023
Accepted: August 18, 2023
Published: September 25, 2023
Identification
D-0087
Citation
Khaliqa Minhas, Muhammad Sufian, Lubna Rasheed & Umer Liaqat. Evaluation of Beauveria Bassiana and Metarhizium Anisopliae in integration with new chemistry insecticide against red cotton Bug (Dysdercus Koenigii) (Fabricius). Dinkum Journal of Natural & Scientific Innovations, 2(09):545-570.
Copyright
© 2023 DJNSI. All rights reserved